|
|
 |
||||||||||||||||
 |
||||||||||||||||
|
Block 1: Ein zusammenfassender Überblick über die Welt der Greifvögel und Eulen Um Wiederholungen bei den einzelnen Abschnitten meiner wissenschaftlichen Ausführungen / Ausarbeitungen zu vermeiden, verweise ich auf das Literaturverzeichnis, das sich auf der Seite “Greifvögel / Eulen 1” befindet. Hiermit untersage ich (Jens Wolf, www kaiseradler.de) jede weitere Verbreitung, Vervielfältigung, Verwertung meiner Fotos in welcher und / oder durch welche Art und / oder Weise auch immer. Genauso untersage ich hiermit jede weitere Verarbeitung, Vervielfältigung, Verwertung meiner wissenschaftlichen Ausführungen / Ausarbeitungen oder Teilen daraus. Das Inhaltsverzeichnis befindet sich auf der Seite “Greifvögel / Eulen A”. Es sei darauf hingewiesen, dass die Möglichkeit besteht, dass im Lauf der Zeit aufgrund von Veränderungen im Bestand der in den Falknereien und/oder (zoologischen) Einrichtungen gehaltenen Arten, die fotografierten Arten nicht mehr gezeigt werden oder nun andere Arten zu sehen sind. |
||||||||||||||||
|
02.01. Die Greifvögel und Eulen, aus der Evolution, vom Beginn und dem Ende der Ersten Die ersten einfachen Landpflanzen entwickelten sich aus den Wasserpflanzen“ vor rund 400 Millionen Jahren, die ersten Bäumen vor rund 300 Millionen Jahren (Der FALKNER 2022, aus Talk Together“; Nr. 74). Aufgrund der starken Abkühlung vor einer Millionen Jahren, bedeckten Eismassen von der Aktis, der Antarktis und den Gebirgen aus grosse Teile Europas, Asiens, Japans und Nordamerikas“ (Der FALKNER 2022, aus Talk Together“; Nr. 74). Nach vier Eiszeiten begannen zuerst Birken und Kiefern, dann auch Eichen, Linden, Ulmen, Eschen und Buchen“ zu wachsen (Der FALKNER 2022, aus Talk Together“; Nr. 74). Im Jahre 1972 verbreitete der britische Paläontologe Dr. Alick D. Walker die These (Norman: Dinosaurier“ (1991)), dass die Vögel aus einem Stamm leicht gebauter Krokodilier-Vorläufer hervorgegangen seien“ (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)). Einige der Krokodilier-Vorläufer begannen nach der Auffassung von Dr. Alick D. Walker auf Bäume zu klettern, andere passten sich der amphibischen Lebensweise an und wurden zu den echten Krokodiliern (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)). Des weiteren besteht die Thesen, dass sich die Vögel von kleinen fleischfressenden Coelurosaurier wie den Dinosaurier Compsognathus“ abstammen oder dass sich die Vögel direkt aus den Vorfahren der Dinosaurier, den Pseudosuchier-Thecodontiern wie Euparkeria“, entwickelten (Lambert: "Alles über das Leben der Vorzeit" (1990)). Inzwischen stimmen jedoch die meisten Experten heute darin überein, dass die Vögel heute möglicherweise von kleinen fleischfressenden Dinosauriern abstammen“ (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)) oder man findet auch, dass die Vorfahren der Vögel“ offenbar Reptilien, die sich in drei große evolutionäre Entwicklungslinien“, den Schildkröten, den Schlangen und Eidechsen und den Krokodilen unterscheiden und dass die heute lebenden Vögel“ zusammen mit den Krokodilen eine Schwestergruppe“ bilden (ORDEN DEUTSCHER FALKONIERE“ (2010)). Es bleiben jedoch die Gemeinsamkeiten aus der Entwicklungsgeschichte der Herrschenden Reptilien, dass Krokodile und Vögel einerseits einen länglichen äusseren Gehörgang, einen Muskelmagen“, vollständig getrennte Herzkammern besitzen und andererseits Nester bauen und eine Brutfürsorge betreiben (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)) oder vollständig getrennte Herzkammern besitzen und an den Hinterfüssen nur vier Zehen“ haben (Schmitt: "Wie sich das Leben entwickelte" (1994)). Als Stammgruppe aller zu den Archosauriern gehörenden Reptilien und“ Vögel gilt wahrscheinlich eine Gruppe der Thecodontier (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)) oder man nimmt an, dass die Vögel sich aus einer Dinosauriergruppe, den Maniraptora entwickelt haben, die ihrerseits eine Untergruppe der Theropoden darstellen, zu der auch bekannte Dinosaurier wie Tyrannosaurus rex gehören (Übersicht in Storch et al., 2007). Während die ursprünglichen Coelurosauria schon einfache Federn aufwiesen, zeigen die Manoraptora bereits ein komplex aufgebautes Federkleid, wie man es bei Archaeopteryx gefunden hat“ (ORDEN DEUTSCHER FALKONIERE“ (2010)). Vor 265 bis 65 Millionen Jahren traten erstmals Krokodile“ auf (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)), vor 249 bis 200 Millionen Jahren wird als ältester Ahn Rauisuchia, ein 4 bis 6m langes Reptil, angegeben (Wunder der Tierwelt“ 05/2016), vor 230 bis 200 Millionen Jahren lebten die Aetosauria, eine 1 bis 3m grosse Reptiliengruppe (Wunder der Tierwelt“ 05/2016), vor 136 bis 65 Millionen Jahren gab es bereits zahlreiche Echte Krokodile“ (Dossenbach: Das Lexikon der Tiere“ (1997)) oder vor 130 bis 65 Millionen Jahren entstanden die Vollkrokodile oder Eusuchier, die zu den heutigen Gruppen führten“ (Lebendige Wildnis, Tiere der Seen und Sümpfe“ (1994)) oder die Linie der Krokodile trat schon vor 240 Millionen Jahren in Erscheinung“ (Spektrum der Wissenschaft: Spezial: Tierische Tricks) oder die Krokodil- und Vogelvorfahren entwickelten vor 300 Millionen Jahren auseinander (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)) oder vor 201 Millionen Jahren trennten sich die Entwicklungslinien der Krokodile auf der einen Seite“ und der Dinosaurier inklusive der Vögel auf der anderen Seite“ (Zoo Dresden: Infotafel). Es werden 22 Krokodilarten in den 3 Familien der echten Krokodile, der Alligatoren und der Gaviale (Lebendige Wildnis, Tiere der Seen und Sümpfe“ (1994)) oder acht Gattungen und 22 Arten“ (Ross/Dr. Garnett: Krokodile und Alligatoren“ (2002)) oder 23 Arten in den 3 Familien der Alligatoren und Kaimane, lat. Alligatoridae mit 4 Gattungen, 8 Arten“, der echten Krokodilen, lat. Crocodylidae mit 3 Gattungen, 12 Arten“ und der Gavialen, lat. Gavialidae mit einer Art (Spektrum der Wissenschaft: Spezial: Tierische Tricks) oder 21 Krokodilarten in den 3 Familien der echten Krokodile, der Alligatoren und Kaimane und der Gaviale (Dossenbach: Das Lexikon der Tiere“ (1997)) oder 24 lebende Arten, die sich in 14 Arten der Echten Krokodile, 8 Arten der Alligatoren und Kaimane und 2 Arten der Gaviale (Zoo Dresden: Infotafel) unterschieden. Genaugenommen zählen zu den echten Krokodilen, lat. Crocodylidae die Gattungen Crocodylus, Osteolaemus, Tomistoma, zu den Alligatoren, lat. Alligatoridae die Gattungen Alligator, Caiman, Melanosuchus, Paleosuchus und zu den Gavialen, lat. Gavialidae die Gattung Gavialis (Grzimeks Tierleben, Enzyklopädie des Tierreichs“, Band 6 (Kriechtiere). |
||||||||||||||||
|
Die nachfolgenden Bilder zeigen Nil-Krokodile, lat. Crocodylus niloticus (aufgenommen im Zoo Köln (links) und im Zoo Halle (Unterart: Crocodylus niloticus cowici; mitte), Leistenkrokodile, lat. Crocodylus porosus (aufgenommen im Zoo Stuttgart (Wilhelma; rechts oben) und Siam-Krokodile, lat. Crocodylus moreletii (aufgenommen in der Welt der Reptilien: Der Zoo“ in Beilrode/OT Döbrichau; rechts unten). |
||||||||||||||||
 |
 |
 |
||||||||||||||
 |
||||||||||||||||
 |
||||||||||||||||
|
Die nachfolgenden Bilder zeigen das Australische Süsswasserkrokodil, lat. Crocodylus johnsoni und dessen Eier (aufgenommen im Zoo Frankfurt (Main; links), ein Sumpfkrokodil, lat. Osteolaemus tetraspis (?; aufgenommen im Zoo Leipzig; mitte), ein Neuguinea-Krokodil, lat. Crocodylus novaeguineae (aufgenommen in der Welt der Reptilien: Der Zoo“ in Beilrode/OT Döbrichau; rechts oben) und ein Beulenkrokodil, lat. Crocodylus moreletii (aufgenommen in der Welt der Reptilien: Der Zoo“ in Beilrode/OT Döbrichau; rechts unten). |
||||||||||||||||
 |
 |
 |
||||||||||||||
 |
||||||||||||||||
 |
 |
|||||||||||||||
|
Die nachfolgenden Bilder zeigen Breitschnautzenkaimane, lat. Caiman latriostris (aufgenommen im Zoo Stuttgart (Wilhelma; links), einen Brillenkaiman, lat. Caiman crocodilus (aufgenommen im Tiergarten Nürnberg; mitte oben; aufgenommen im Zoo Rostock; mitte unten), einen Brauen-Glattstirnkaiman, lat Paleosuchus palpebrosus (aufgenommen im Tierpark Chemnitz; rechts oben) und einen Geilkopf-Glattstirnkaiman, lat Paleosuchus trigonatus (aufgenommen in der Welt der Reptilien: Der Zoo“ in Beilrode/OT Döbrichau; rechts unten). |
||||||||||||||||
 |
 |
 |
||||||||||||||
 |
||||||||||||||||
 |
 |
|||||||||||||||
|
Die nachfolgenden Bilder zeigen einen Hechtalligator, lat. Alligator mississippiensis (aufgenommen im Tiergarten Berlin (Friedrichsfelde); links), einen China-Alligator, lat. Alligator sinensis (aufgenommen im Tiergarten Berlin (Friedrichsfelde); mitte) und einen Sunda-Gavial, lat. Tomistoma schlegeli (aufgenommen im Zoo Dresden: rechts). |
||||||||||||||||
 |
 |
 |
||||||||||||||
 |
 |
|||||||||||||||
|
Vor 220 Millionen Jahren lebten in Zentralasien Reptilien (Longisquama insignis), die Federn besessen haben sollen (FALKE 10/2000). Andererseits findet man auch, dass die Entwicklung der Federn vor rund 200 Millionen Jahren“ begann und die Federn wohl zunächst vor allem der Isolation zum Schutz vor Kälte beziehungsweise dem konstant halten der Körpertemperatur“ dienten (ORDEN DEUTSCHER FALKONIERE“ (2018)). Die Entwicklung der Federn führte später auch zu hervorragenden Flugeigenschaften und letztendlich zum aktiven Flug der Vögel“ (ORDEN DEUTSCHER FALKONIERE“ (2018)). In der Mongolei (Falke 12/2008) wurde ein ca. 150 Millionen Jahre alter nicht flugfähiger mit Federn bedeckter Dinosaurier (Epidexipteryx hui aus der Gruppe Theropoda) mit 4 langen Schwanzfedern (zur Balz benutzend?) gefunden. Forscher der Chinesischen Akademie der Wissenschaften (FALKE 06/2010) fanden beim ältesten Federsaurier Sinosauropteryx Überreste von Pigmenten, die Rückschlüsse auf Farbe und Federkleidzusammensetzung zulassen. Des Weiteren wird die andererseits Annahme (FALKE 06/2010) unterstützt, da die Flaumfedern nicht den gesamten Körper bedeckten, dass die Federn Signalfunktion, jedoch keine Kälteschutzfunktion hatten und auch nicht zum fliegen verwendet wurden. Man findet, dass die hypothetische bisher noch nicht gefundene Urvogelform“ Proavis im Zeitraum vor etwa 250 Millionen Jahren“ einzuordnen ist und zu Beginn der Entwicklungslinie der Vögel bei der ausgestorbenen kleinen Reptilien-Gruppe Pseudosuchia zu suchen sei“ (Benes/Burian: Tiere der Urzeit“ (1980)), dass Proavis, der hypothetische Vorvogel zeitlich vor vor 200 Mill. Jahren eingeordnet” wird (Deutschen Greifenwarte Bernolph Frhr. von Gemmingen, Burg Guttenberg (früher: Deutsche Greifenwarte Claus Fentzloff, Burg Guttenberg: Hinweis), dass der hypothetische Vor-Vogel“ Proavis war ein zweifüssiges rennendes Reptil, dessen Vordergliedmassen zunächst nur luftrudernd gebraucht wurden“ und später zu Flügeln wurden, mit denen sich das Tier in die Luft schwingen konnte“ (Krösche: Die Moa-Strausse“ (2006)), dass der Proavis, die Vorstufe des Archaeopteryx“, war nach einer Hypothese ein arborealer primitiver Klammerkletterer, der den Gleitflug erwarb“ oder ein zweifüssiger Renner, der mit den Vorderextremitäten lebende Beute niederschlug“ (Kuhn-Schnyder, Rieber: "Paläozoologie" (1984)), dass Überreste einer solchen Übergangsform zwar nicht entdeckt worden“ sind, aber man hat diese als Proavis bezeichnet“ (Krösche: Die Moa-Strausse“ (2006)). |
||||||||||||||||
|
|

|
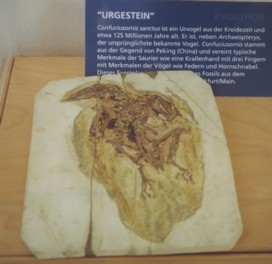
Zum Protoavis findet man, dass Protoavis im Zeitalter Trias lebte, eine Grösse von etwa 60cm und eine Flügelspannweite von 70cm hatte (Czerkas/Czerkas: Dinosaurier“ (1990)), dass ein kleiner, vogelähnlicher Fleischfresser“ in Texas gefunden wurde, der im Zeitalter Oberes Trias lebte und als Protoavis 1991 beschrieben wurde (Lambert: "Dinosaurier" (2002)), dass Dr. Sankar Chatterjee von der texanischen Universität in Lubbok den Protoavis entdeckte, auch eine Pressemeldung veröffentlichte (Norman: Dinosaurier“ (1991), Seiten 137, 142), dass im damaligen Tropenwald von Texas“ der krähengrosse Ururvogel (Protoavis texensis)“ lebte und vor etwa 215 Millionen Jahren“ ausstarb (Urania Tierreich, Band Vögel“ (2000)) oder dass Protoavis texensis langschwanzig war, jedoch Federn fand man bei Protoavis nicht“ und 75 Millionen Jahre älter als Archaeopleryx“ ist (Anzeiger des Vereins Thüringer Ornithologen“: 2006: Heft 3) oder dass vor rund 225 Millionen Jahren in Texas Ablagerungen des etwa fasanengrossen Protoavis“ gefunden wurden, jedoch noch immer noch nicht ausreichend dokumentiert bzw. publiziert wurden (FALKE 09/1995), dass zum Erstvogel“, lat Protoavis, bislang nicht zweifelsfrei geklärt werden“ konnte, ob Protoavis überhaupt ein Vogel war“, weil die Knochen derart unvollständig erhalten“ geblieben sind und die Federn und andere wesentliche Merkmale eines fliegenden Vogels“ fehlen, so dass man zur Schlussfolgerung kommt, dass Protoavis vermutlich kein Vogel, sondern ein am Boden lebendes Reptil“ war (FALKE 07/1997). Der Urvogel, lat. Archaeopteryx hatte die Grösse eine Elster“ ("Das grosse Buch des Allgemeinwissens", 1992; Perrins: Vögel“) oder war nicht grösser als eine Elster“ (Ackroyd: "Die Entstehung des Lebens, die Geschichte der Welt“ (2004)) oder war etwa elsterngross (Buch der Vogelwelt, Mitteleuropas“ (1973)) oder war rabengross (NABU: Naturschutz-heute“ (01/2010)) oder hatte etwa die Grösse einer Krähe oder einer Taube“ (Forshaw: "Enzyklopädie der Tierwelt", Band Vögel (1991)) oder war ca. 30cm gross (Richardson: "Dinosaurier und andere Tiere der Urzeit" (2004)) oder war 35cm gross (Benton: Tiere der Vorzeit von A-Z“ (1991)) oder war 40cm gross (Archaeopteryx lithographica: Rheinwald: "Atlas der Vogelwelt" (1994)), lebte in Europa (Forshaw: "Enzyklopädie der Tierwelt", Band Vögel (1991)), oder lebte an den Gestaden eines Meeres in Deutschland“ (Zillmer: "Dinosaurier-Handbuch" (2002)) oder lebte am Seeufer und den offenen Waldlandschaften (Richardson: "Dinosaurier und andere Tiere der Urzeit" (2004)) im Zeitraum von vor etwas mehr als 146“ Millionen von Jahren (1000 Fragen, 1000 Antworten: Wunderbare Tierwelt“, Readers Digest Wissenswelt (2006)) oder vor 200 bis 145 Millionen Jahren (Forshaw: "Enzyklopädie der Tierwelt", Band Vögel (1991)) oder vor etwa 160 Millionen Jahren“ (Everett: Raubvögel der Welt“ (1978)) oder vor ca. 150 Millionen von Jahren (Benes/Burian: Tiere der Urzeit“ (1980); Ivanov, Hrdolickova, Gregorova: Illustrierte Fossilien Enzyklopädie“; Ackroyd: "Die Entstehung des Lebens, die Geschichte der Welt“ (2004); Rheinwald: Atlas der Vogelwelt“ (1994); Buch der Vogelwelt, Mitteleuropas“ (1973); Benton: Tiere der Vorzeit von A-Z“ (1991); Greifvögel und Falknerei“ (1989); NABU: Naturschutz-heute“ (01/2010); Unwin: Atlas der Vögel“ (2012); Anzeiger des Vereins Thüringer Ornithologen“: 2006: Heft 3) oder vor 156 bis 150 Millionen von Jahren (Zillmer: "Dinosaurier-Handbuch" (2002); Lambert: "Dinosaurier" (2002)) oder vor 130 Millionen Jahren (Diesselhorst, Fechter: Knaurs Tierleben von A-Z, Band 2 (M-Z)”, 1981) oder vor 140 Millionen von Jahren (Nachtigal: Vogelflug und Vogelzug“ (1987); Fisher, Peterson: "Das Bunte Buch der Vögel" (1979); "Zum Fliegen geboren; Das Weltreich der Vögel" (1988); Archaeopteryx lithographica: Urania Tierreich, Band Vögel“ (2000)) oder im Jura vor ungefähr 140 Millionen Jahren“ (Perrins: Vögel“) oder im Jura vor ungefähr 140 Millionen Jahren“ (Perrins: Vögel“) oder vor ca. 147 Mio. Jahren“ (Perrins: "Die BLV Enzyklopädie, Vögel der Welt", 2004). Archaeopteryx heisst übersetzt griech. Archaios, >uralt< und pteryx, >Flügel<“ ("GEOkompakt: Die Urzeit" Nr. 8) oder sehr alter Flügel“ (Lambert: "Dinosaurier" (2002)) oder alter Flügel“ (Lambert: "Alles über das Leben der Vorzeit" (1990); FALKE 07/1997) oder alte Feder“ (NABU: Naturschutz-heute“ (01/2010)). Die erste versteinerte 8cm lange Feder fand man im Jahre 1860 (Urania Tierreich, Band Vögel“ (2000)) oder man fand als ersten Fund den Abdruck einer Feder“ eines Archaeopteryx (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005)) oder man fand bereits 1860 eine einzelne Feder“, die einen Hinweis auf die Existenz von Vögeln in der Jurazeit“ lieferte (FALKE 07/1997) oder man fand eine etwa 7cm“ lange versteinerte Feder im Fühling oder Frühsommer 1861“ (FALKE 08/2024), die von Hermann von Meyer im Jahre 1861 als Archaeopteryx lithographica beschrieb“ (Kuhn: "Die vorzeitlichen Vögel" (1971)) oder die den Namen Archaeopteryx lithographica von Meyer im Jahr 1861 bekam (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979)). Im Jahr 1861 entdeckte man ein Skelett, einen Archaeopteryx, der an das Britische Museum in London verkauft wurde (Werner: 1000 Dinosaurier“; Norman: Dinosaurier“ (1991)) oder das 1861 gefundene Skelett wird als Londoner Exemplar bezeichnet (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005), FALKE 08/2024). Owen beschrieb im Jahr 1864 das Skelett mit dem Namen Archaeopteryx macrura (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979)) oder man findet aber auch, dass das erste Skelettexemplar im Herbst 1862 nach London“ kam und im Jahr 1883“ mit dem Namen Archaeopteryx macrura bechrieben wurde (FALKE 2024). Im Jahr 2011 entschied nun die Internationale Kommission für Zoologische Nomenklatur ICZN, dass das Londoner Urvogelexemplar zum Neotypus von Archaeopteryx lithographica v. Meyer, 1861“ wird und damit hat die Urvogel-Feder ihren Status als Holotypus verloren“ und ist seitdem formal ohne einen wissenschaftlichen Namen und sozusagen >namenslos< geworden“ (FALKE 08/2024). Im Jahr 1877 entdeckte man Archaeopteryx siemensi, der an das Berliner Museum für Naturkunde verkauft wurde (Werner: 1000 Dinosaurier“), das sogenannte Berliner Exemplar ist der dritte Fund (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005); Norman: Dinosaurier“ (1991), FALKE 08/2024). Archaeopteryx siemensi ist kleiner und entweder ein nicht voll erwachsener Archaeopteryx oder ein Weibchen (Werner: 1000 Dinosaurier“). Den Namen Archaeopteryx siemensi bekam das Berliner Exemplar von Dames im Jahr 1897 (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979)), ursprünglich wählte man den Namen Archaeornis siemensi, weil man diesen Fund für eine andere Art und Gattung hielt (Urania Tierreich, Band Vögel“ (2000)). In FALKE (08/2024) findet man nur die Bezeichnung Berliner Exemplar, lat. Archaeopteryx und als Jahr der Erstbeschreibung 1875(!?). Im Jahr 1956 entdeckte man als vierten Fund das Maxberger Exemplar (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005), FALKE 08/2024). Das Maxberger Exemplar ist stark zerfallen und man findet man nur die Bezeichnung Maxberger Exemplar, lat. Archaeopteryx (FALKE 08/2024). Im Jahr 1957 entdeckte man als fünften Fund ein Exemplar, welches sich im Teyler Museum, Haarlem befindet (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005)). Zum Haarlemer Exemplar findet man die Bezeichnung Urvogel Ostromia crassipes, 1855“ und dass Haarlemer Exemplar ist unvollständig (FALKE 08/2024). Im Jahr 1951 entdeckte man als sechsten Fund das Eichstädter Exemplar (Stephan: "Urvögel", NBB 465, 2.Aufl. (1979), 4.Aufl. (2005), FALKE 08/2024). Das Eichstädter Exemplar ist nahezu vollständig gut erhalten und man findet man nur die Bezeichnung Eichstädter Exemplar, lat. Archaeopteryx (FALKE 08/2024). Ivanov, Hrdolickova, Gregorova (Illustrierte Fossilien Enzyklopädie“), Kuhn-Schnyder, Rieber ("Paläozoologie" (1984)) und FALKE (09/1995) geben an, dass bereits 7 Funde des Archaeopteryx beschrieben wurden. Mit der Angabe vor 1985“ wird das grösster Exemplar, das Solnhofener Exemplar mit dem Namen Solnhofener Exemplar, lat. Archaeopteryx beschrieben (FALKE 08/2024. Im Jahr 1992 wird das weitgehend vollständig erhaltene Münchener Exemplar beschrieben und man findet nur nur die Bezeichnung Münchener Exemplar, lat. Archaeopteryx (FALKE 08/2024). Um das Jahr 1990 wird das unvollständig erhaltene Daitinger Exemplar beschrieben und man findet nur nur die Bezeichnung Daitinger Exemplar, lat. Archaeopteryx (FALKE 08/2024). Mit dem Jahr 2004 wird ein Flügelrest, das Exemplar der Familien Ottmann & Steil mit der Bezeichnung Exemplar der Familien Ottmann & Steil, lat. Archaeopteryx (FALKE 08/2024) angegeben. In FALKE (03/2006, 02/2007) wird angegeben, dass man ein zehntes Exemplar des Archaeopteryx untersuchen konnte, welches in schweizer Privatbesitz gewesen ist und dass keinerlei Angaben zum wer“ und wann“ das zehnte Exemplar fand, gemacht werden konnten. Das 10. Exemplar, das Thermopolis-Exemplar, ist vollständig und ausgezeichnet erhalten“ und man findet nur nur die Bezeichnung Thermopolis-Exemplar, lat. Archaeopteryx (FALKE 08/2024). Im Jahr 2014 wurde das vollständig und ausgezeichnet mit Federkleid“ erhaltene Altmühltal-Exemplar mit dem Namen Altmühltal-Exemplar, lat. Archaeopteryx beschrieben (FALKE 08/2024). Im Jahr 2010 wurde das älteste vollständig erhaltene Schamhauptener-Exemplar mit dem Namen Schamhauptener-Exemplar, Archaeopteryx beschrieben (FALKE 08/2024). Im Jahr 2017 wurde der unvollständige Urvogel Alcmonavis poeschli beschrieben (FALKE 08/2024). Im Jahr 2019 wurde ein weiterer unvollständiger Urvogel entdeckt, der derzeit in wissenschaftlicher Bearbeitung“ ist (FALKE 08/2024). Im Jahr 2024 wurde vom Field Museum in Chicago, USA“ ein nahezu vollständig und ausgezeichnet mit Federkleid“ erhaltener Fund, höchstwahrscheinlich Archaeopteryx“, derzeit in wissenschaftlicher Bearbeitung“ bekanntgegeben (FALKE 08/2024). |